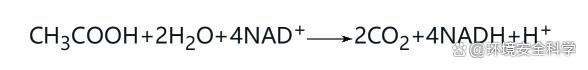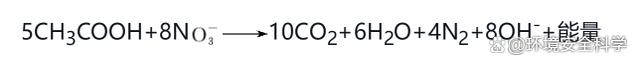當前位置:首頁>生活> 污泥處理的厭氧微生物的產能代謝
發布時間:2025-10-28閱讀( 15)
|
厭氧微生物的產能代謝主要包括兩種類型,即發酵和無氧呼吸。
(1)發酵 發酵(fermentation)是某些厭氧微生物在生長過程中獲得能量的一種方式,以有機物氧化分解的中間代謝產物為最終電子受體的氧化還原過程。在發酵過程中,可被利用的底物通常為單糖或某些雙糖,亦可為氨基酸等。其最終產物是有機酸、醇、CO2、H2及能量。 1)糖酵解途徑 (www.ws46.cOm) 糖酵解(embdem-meyerhof-parnas,EMP)主要分為2個步驟。步驟1包括一系列不涉及氧化還原反應的預備性反應,主要是通過加入能量使葡萄糖活化,并將六碳糖分解為三碳糖,其結果是生成一種主要的中間產物3-磷酸甘油醛,并消耗2molATP;步驟2是通過氧化還原反應,產生4molATP、2molNADH H 和2mol丙酮酸。 糖酵解途徑具有以下重要意義。 Ⅰ.其產能方式稱為底物水平磷酸化,分別發生在1,3-二磷酸甘油酸與2-磷酸烯醇式丙酮酸兩處。所謂底物水平磷酸化是指底物被氧化過程中,在中間代謝產物分子上直接形成比高能焦磷酸鍵含能更高的高能鍵,并可直接將鍵能交給ADP使之磷酸化,生成ATP的這一過程。底物水平磷酸化是進行發酵的微生物獲取能量的唯一方式。1mol葡萄糖經酵解后凈產2molATP。 Ⅱ.微生物通過發酵可將葡萄糖轉變成乳酸、丙酸、乙醇、丁醇等多種代謝產物,糖酵解是產能的主要途徑,產生的能量作為各種發酵產物產生的主要甚至唯一的能量來源。 ② 丙酮酸是重要的中間代謝物
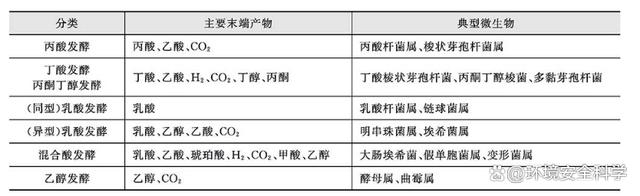
糖酵解的終產物是丙酮酸,通過丙酮酸的進一步發酵,可產生各種發酵產物,并可通過NADH H 的氧化,使機體內NADH H 含量保持在一定范圍內,從而保證發酵的正常進行。 2)主要發酵類型 微生物發酵形式多樣,普遍存在的是以糖酵解為主體的分解代謝,其末端產物則各不相同,發酵類型是根據主要末端產物命名的。微生物機體內調控某一發酵類型中末端產物種類及數量的原因主要有兩點。其一,氧化反應必須與另一個還原反應相偶聯,從而維持NADH H /NAD 在一定范圍內,這種偶聯是通過輔酶在兩者之間反復地還原和氧化,不斷周轉而完成的;其二,發酵產能是微生物的目的所在,微生物可根據能量需求狀況來調整高產能發酵產物的轉化率。表4-5為碳水化合物發酵的主要類型。
發酵在廢水和污泥厭氧消化過程中起著非常重要的作用,甚至此過程比產甲烷過程更為關鍵。國內外的研究表明,在厭氧消化過程中主要存在丙酸型發酵和丁酸型發酵兩種類型。這兩種發酵類型的分類與生物化學中的分類有一定聯系,但也有所差別。 ① 丙酸發酵。 丙酸發酵以糖酵解產生的丙酮酸為起點,其中包括部分TCA循環。主要參與的細菌是丙酸桿菌屬。此類型發酵的特點是氣體(CO2)產量很少,甚至有時無氣體產生,主要發酵末端產物為丙酸和乙酸。總反應式如式所示:
在廢水厭氧處理過程中,含氮有機化合物(如明膠、酵母膏、肉膏等)和難降解碳水化合物(如纖維素)常呈丙酸型發酵(末端產物與丙酸型發酵類似)。丙酸型發酵在厭氧生物處理過程中不夠理想,這是因為末端產物丙酸不易轉化為可被產甲烷菌利用的底物,易出現丙酸積累。當丙酸出現大量的積累,會導致厭氧反應器內的pH值的大幅度下降,從而因產甲烷菌失去活性而導致厭氧反應器的運行失敗。 ② 丁酸發酵。 丁酸的循環機制起著重要作用,一方面使糖酵解途徑及產乙酸過程中釋放的NADH H 通過與丁酸產生相偶聯而得以氧化;另一方面還可以減少酸性末端。解糖梭狀芽孢桿菌進行此類反應。
盡管如此,當pH值降至4.5以下時,丁酸循環機制被阻斷,轉為形成中性末端產物的丙酮丁醇發酵。 該發酵類型與廢水厭氧生物處理過程中的丁酸型發酵類似,許多研究結果表明,含可溶性碳水化合物(如葡萄糖、蔗糖、淀粉、乳糖等)廢水的發酵常出現丁酸型發酵,發酵中主要的末端產物為丁酸、乙酸、二氧化碳、氫和及少量丙酸。當運行管理不當時,丙酸含量顯著增加,甚至有可能轉化為丙酸型發酵。 ③ 混合酸發酵。 由于在發酵產物中存在許多有機酸,因此稱為混合酸發酵。進行該類型發酵的主要是一些腸道細菌,如志賀菌屬、埃希菌屬等。這些細菌是一些兼性厭氧菌,在有氧情況下進行呼吸,在缺氧情況下進行發酵。 埃希菌由于存在甲酸氫解酶,可將甲酸分解為氫和二氧化碳,因此,大腸桿菌可以產氣,而志賀菌等由于不存在甲酸氫解酶,故不能將甲酸分解為氫和二氧化碳,因此不產氣。可以通過產酸產氣試驗將一些不產酸以及產酸不產氣的細菌區分開來。 有些細菌如產氣腸桿菌在發酵時,除了將一部分丙酮酸按混合酸發酵的類型進行外,大部分丙酮酸先通過兩個分子的縮合成為乙酰乳酸,再脫羧為3-羥基丁酮,然后再還原為丁二醇。 3-羥基丁酮在堿性條件下被空氣中的氧氣氧化為乙二酰。根據乙二酰能與胍基作用生成紅色化合物的特點,可以測定3-羥基丁酮的存在,這就是V.P.試驗為陰性,而產氣腸桿菌V.P.試驗為陽性。此外,大腸桿菌由于產酸較多,所以pH值低于4.5,用甲基紅作指示劑可顯出,而產氣腸桿菌的產物丁二醇是中性化合物,因此通過V.P.試驗和甲基紅試驗可對兩種細菌進行鑒別。
④ 乙酸發酵 進行乙酸發酵的微生物主要是酵母菌,如釀酒酵母,其發酵葡萄糖的末端產物僅有乙醇和CO2。 有些厭氧細菌亦可進行乙酸發酵,但其代謝途徑和酵母菌不同。例如螺旋體屬發酵碳水化合物,在代謝中與梭狀芽孢桿菌屬類似。發酵葡萄糖生成的主要末端產物為乙酸、乙醇、氫和二氧化碳。螺旋體屬為專性或兼性厭氧微生物,存在于廢水和活性污泥中。 乙醇型發酵與細菌性乙醇發酵相似,主要終產物為乙酸、乙醇、丁酸、氫、二氧化碳及極少量的丙酸。
乙醇型發酵的末端產物極為理想,丙酸產物很少,且乙醇很容易轉化為產甲烷菌可利用的底物(乙酸、CO2和H2)。
(2)無氧呼吸 進行無氧呼吸的厭氧微生物生活在河、湖、池塘底部淤泥等缺氧環境中,以硝酸根、碳酸根、硫酸根等作為最終電子受體的氧化還原過程,進行有機物的生物氧化。最終產物是N2、H2S、CH4、CO2、H2O及能量。 1)硝酸鹽呼吸 硝酸鹽呼吸也稱異化型硝酸鹽還原。缺氧條件下,有些細菌能以有機物為供氫體,以硝酸鹽作為最終電子受體,這類細菌稱為硝酸鹽還原菌。不同的硝酸鹽還原菌將硝酸根還原的末端產物不同,如N2(包括N2O、NO)、NH3和亞硝酸根。 通過硝酸鹽呼吸將硝酸根還原為氣態N2(包括N2O、NO)的過程稱為反硝化作用。能夠進行反硝化作用的細菌稱為反硝化細菌,主要有反硝化假單胞菌、銅綠假單胞菌、施氏假單胞菌、地衣芽孢桿菌、反硝化副球菌等,其中某些菌可兼性好氧。這些細菌可將有機底物徹底氧化為CO2,同時伴隨脫氫反應,如式所示:
NADH H 經電子傳遞體系將最終電子受體N[插圖]還原為N2,同時伴隨能量的產生。
亞硝酸對細菌來說是毒性物質,因此它的積累不利于細菌生長。對于大多數細菌來說,亞硝酸鹽還原酶是一個誘導酶,亞硝酸鹽的產生將誘導產生亞硝酸鹽還原酶,并迅速將亞硝酸鹽還原產生末端產物,如反硝化副球菌將亞硝酸根轉化為N2;而大腸埃希菌將硝酸根轉化為NH3。對于無亞硝酸鹽還原酶合成機制的硝酸鹽還原細菌則只能在有限的亞硝酸鹽濃度范圍內利用硝酸鹽。 硝酸鹽還原作用給農業生產帶來較大的損失。一般情況下,施入水稻田里的氮肥由于硝酸鹽還原作用而損失1/2。但是,從自然界物質循環角度考慮,硝酸鹽還原作用是有利的。在廢水處理中,為降低水中含氮量所采取的生物脫氮法就是基于反硝化作用原理。
2)硫酸鹽呼吸 硫酸鹽呼吸常稱為異化型硫酸鹽還原或反硫化作用。參與細菌主要為無芽孢的脫硫弧菌屬和形成芽孢的脫硫腸狀菌屬,均為專性厭氧、化能異養型細菌。大多數硫酸鹽還原菌不能利用葡萄糖作為能源,而是利用丙酮酸和乳酸等其他細菌的發酵產物。乳酸和丙酮酸等作為供氫(電子)體,經無NAD 參與的電子傳遞體系將硫酸根還原為H2S。 3)碳酸鹽呼吸 碳酸鹽呼吸即異化型碳酸鹽還原,亦可稱作產甲烷作用,過去人們常誤稱作甲烷發酵。進行碳酸鹽還原的細菌稱為產甲烷細菌。 產甲烷菌專性厭氧,僅能以甲酸、甲醇、甲胺、乙酸和H2/CO2作為底物(供氫體)。產甲烷菌不含N-乙酰胞壁酸和N-乙酰葡萄糖胺組成的肽聚糖,對青霉素(可阻止細胞壁合成)不敏感。因而在產甲烷菌的分離中,可利用這一特性抑制非產甲烷菌的生長,達到分離產甲烷菌的目的。 產甲烷菌除存在于缺氧的沼澤地以及河、湖、池塘的淤泥中外,在反芻動物的瘤胃中也含有。產甲烷菌在廢水、污泥厭氧生物處理中起重要作用,底物經發酵細菌轉化為乙酸、甲酸、甲醇、甲胺及H2/CO2后,在產甲烷菌的厭氧呼吸下生成CH4和CO2。在廢水厭氧生物處理中常見的產甲烷菌有產甲烷八疊球菌屬、產甲烷桿菌屬、產甲烷短桿菌屬、產甲烷球菌屬、產甲烷螺菌屬及產甲烷絲菌屬等。 |
Copyright ? 2024 有趣生活 All Rights Reserve吉ICP備19000289號-5 TXT地圖HTML地圖XML地圖